




Figures
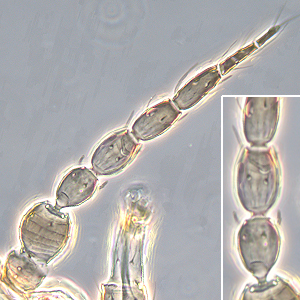
Fig. 1: Antenna (inset: III. and IV. antennal segment)
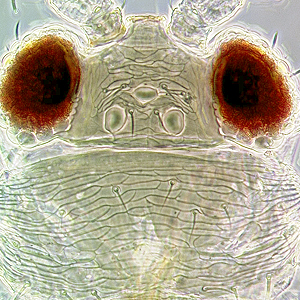
Fig. 2: Head dorsal with ocellar triangle
Fig. 3: Pronotum
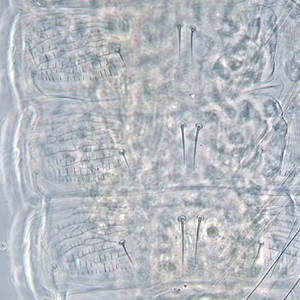
Fig. 4: Meso- and metanotum
Fig. 5: Fore- and hindwing, base of fore wing with alula
Fig. 6: Metathoracic furca
Fig. 7: Tergites V - VII
Fig. 8: Tergites VII and VIII
Taxonomic Information
Species:
Pseudodendrothrips mori (Niwa, 1908)
Synonyms:
Graphidothrips stuardoi Moulton, 1930
Belothrips
mori Niwa, 1908
Common name:
Mulberry thrips
Present taxonomic position:
Family: Thripidae Stephens, 1829
Subfamily: Dendrothripinae Priesner, 1925
Genus: Pseudodendrothrips Schmutz, 1913
Species Recognition
General information about the genus Pseudodendrothrips:There are currently 16 species recognized in the genus Pseudodendrothrips and are distinguished from other genera in having the eyes bulging from the side of the head and the head inset: between the eyes, the antennae are eight segmented, segment II wider than the rest and with forked sense cones on segments III and IV, and the pronotum contains two long setae. The species in this genus also have a lyre-shaped metathoracic endofurca present, the forewing has only one or two setae on the first vein and none on the second and in addition there a postmarginal comb present on tergite VIII.
Typical character states of Pseudodendrothrips mori:
Body color
Mainly pale or yellow, with some darker markings
Antennae
Number of antennal segments: 8-9
Segment IV - forked sensorium: scarcely extending beyond base of segment V
Segments II and III shape: more or less symmetric
Segments III & IV sensoria: emergent and forked
Base of sensorium on antennal segment VI: no more than 2 times as wide as base of nearest seta
Terminal antennal segments: rarely elongate
Head:
Distance between bases of ocellar setae III: greater than width of first ocellus
Head shape between compound eyes: not prolonged
Ocellar setae III on head: arising between hind ocelli, or behind tangent of anterior margin of hind ocelli or within ocellar triangle anterior to tangent of anterior margin of hind ocelli
Postocular setae I: present
Surface of head, pronotum and fore legs: without strong reticulate sculpture
Ocellar setae I in front of anterior ocellus: present
Prothorax
Number of pairs of elongate pronotal setae: 0-3
Number of pairs of elongate posteroangular pronotal setae: 1
Pronotum shape: rectangular
Mesothorax
Mesothoracic endofurca: without median spinula
Metathorax
Metanotal median area sculptured lines: transverse at anterior, but longitudinal and parallel on posterior half
Metanotal median setae length: shorter than lateral metanotal setae
Metanotal median setae position: arising behind anterior margin
Metanotum: with campaniform sensilla
Metanotum major sclerite: with two major sclerites, metascutum and metascutellum
Metanotum median area: with no equiangular reticulation
Metanotum sculpture: without dominant sculptured triangle medially
Metathoracic endofurca: elongate and lyre-shape
Wings
Wings: present and more than half as long as abdomen
First vein of forewing: distinct from costal vein
Forewing anterior margin: with setae and cilia but cilia longer than setae
Forewing color: uniformly pale or weakly shaded
Forewing costal fringe of cilia: arising at anterior margin of wing
Forewing costal setae at middle of wing: shorter than median width of wing
Forewing first vein setal row: incomplete, with setae not closely and uniformly spaced
Forewing posterior margin cilia: straight, no undulations
Forewing second vein setal row: with no setae
Forewing surface: not reticulate
Forewings: with veins, setae and microtrichia
Legs
Fore tibial apex: not extending around fore tarsus
Mid and hind tarsi: with one segment
Abdomen:
Abdominal segment X: never tubular, longitudinally incomplete ventrally in both sexes
Abdominal sternite III of female: without glandular areas
Abdominal sternite VII: with marginal setae but no discal setae
Abdominal sternite VII median marginal setae: arising at margin
Abdominal sternites IV , V and VI: with marginal setae but no discal setae
Abdominal tergites: without curved wing-retaining setae
Abdominal tergites IV & V median setal pair: longer than distance between their bases
Abdominal tergites V-VII: without paired ctenidia, sometimes with irregular microtrichia
Setae on abdominal tergite X: slender
Surface of lateral thirds of abdominal tergites: with many microtrichia-like longitudinal ridges
Ctenidia on tergite VIII: not present, but groups of microtrichia
Tergite VIII posteromarginal comb of microtrichia: present, complete medially
Tergite VIII posteromarginal microtrichia: long, slender and regular
Biology
Life history:
As with other thrips species the life cycle from egg to adult is dependent on temperature. The full cycle can take about 15 days to over a month and adults may live for more than one month producing several generations in one year depending on seasonal weather (Lewis,
1973). With greenhouse temperatures the developmental time from egg to adult can decrease to about one week.
Host plants:
Morus, Ficus
Vector capacity:
None identified
Current known distribution:
Africa, Asia, Australia, New Zealand, Central and South America, Europe, North America
Additional notes:
Living on the underside of newly emerged
leaves of their plant hosts and actively disperse when disturbed.
Bibliography
Kakimoto, K, Inoue, H, Hinomoto, N, Noda,
T, Hirano, K, Kashio, T, Kusigemati, K &
Okajima, S (2006): Potential of Haplothrips brevitubus (Karny)
(Thysanoptera : Phlaeothripidae) as a predator of mulberry thrips Pseudodendrothrips
mori (Niwa) (Thysanoptera : Thripidae). - Biological Control 37 (3):
314-319.
Kakimoto, K, Inoue, H & Yoshida, T (2002): Seasonal
changes in population density and species composition of Orius spp.
in mulberry fields. - Japanese Journal of Applied Entomology and Zoology
46 (4): 209-215.
Lewis, T (1973): Thrips their biology,
ecology and economic importance. Academic Press Inc., London Ltd. 349
pp.
Miyazaki, M & Kudo, I (1989): Descriptions
of thrips larvae which are noteworthy on cultivated plants (Thysanoptera)
.2. Pseudodendrothrips-Mori (Niwa). - Applied Entomology and Zoology
24 (2): 209-212.
Moritz G, Morris DC, Mound LA (2001): ThripsID -
Pest thrips of the world. ACIAR
and CSIRO Publishing Collingwood, Victoria, Australia, CDROM ISBN
1 86320 296 X.
Moritz G, Mound LA, Morris DC, Goldarazena A (2004): Pest
thrips of the world - an identification and information system using molecular
and microscopial methods. CBIT, University of Queensland,CDROM ISBN 1-86499-781-8.
Mound,
LA (1999): Saltatorial leaf-feeding Thysanoptera (Thripidae : Dendrothripinae)
from Australia and New Caledonia, with newly recorded pests of ferns, figs and
mulberries. - Australian Journal of Entomology 38: 257-273.
Mound, LA & Kibby, G (1998): Thysanoptera: An identification
guide, (2nd edition). CAB International, Wallingford and New York,
70pp.
Mound, LA & Marullo, R (1996): The
thrips of Central and South America: An Introduction (Insecta: Thysanoptera).
Associated Publishers, Gainesville.
Palmer, JM, Mound, LA & Du Heaume, GJ (1989): 2. Thysanoptera,
pp. 73. In Betts, CR [ed.], CIE Guides to Insects
of Important to Man. CAB International, Wallingford.
Stannard, LJ (1968): The thrips, or Thysanoptera, of Illinois.
Illinois Natural History Survey Bulletin 29: 215-552.
Links:
Mound, LA (2005): Thysanoptera (Thrips) of the World
- A Checklist. http://www.ento.csiro.au/thysanoptera/worldthrips.html